Trao đổi chất – Wikipedia tiếng Việt
Trao đổi chất (còn gọi là chuyển hóa, biến dưỡng, metabolism trong tiếng Anh, lấy từ tiếng Hy Lạp: μεταβολή metabolē, “biến đổi”) là tập hợp các biến đổi hóa học giúp duy trì sự sống trong các tế bào của sinh vật. Ba mục đích chính của quá trình trao đổi chất là chuyển đổi thức ăn/nhiên liệu thành năng lượng để sử dụng cho các quá trình của tế bào, biến đổi thức ăn/nhiên liệu thành các đơn vị để tạo nên protein, lipid, axit nucleic cùng một số carbohydrate và loại bỏ chất thải chuyển hóa. Những phản ứng này được xúc tác bởi các enzym cho phép các sinh vật sinh trưởng và sinh sản, duy trì cấu trúc bản thân và đáp ứng với môi trường xung quanh. Thuật ngữ “trao đổi chất” cũng có thể dùng để chỉ tất cả các phản ứng hóa học xảy ra trong sinh vật sống, bao gồm tiêu hóa và vận chuyển các chất giữa các tế bào hoặc giữa tế bào với môi trường, trong trường hợp các phản ứng diễn ra trong tế bào thì được gọi là chuyển hóa trung gian hoặc trao đổi chất trung gian.
Trao đổi chất thường được chia thành hai loại chính : dị hóa, quy trình ” phá vỡ ” những chất hữu cơ ví dụ như, phân giải glucose thành pyruvate trong hô hấp tế bào ; và đồng điệu, quy trình ” thiết kế xây dựng ” những thành phần của tế bào như protein và axit nucleic. Thông thường, dị hóa sẽ giải phóng năng lượng và đồng nhất thì tiêu tốn năng lượng .Các phản ứng hóa học trong trao đổi chất được tổ chức triển khai thành những con đường chuyển hóa, trong đó một chất hóa học được đổi khác trải qua một loạt những bước để thành một chất khác, với sự tham gia của một chuỗi những enzym. Enzym rất quan trọng trong trao đổi chất do tại những phân tử này được cho phép những sinh vật đẩy nhanh vận tốc những phản ứng yên cầu năng lượng bằng cách kết cặp chúng với những phản ứng tự phát giải phóng năng lượng. Nếu không có enzym, những phản ứng yên cầu năng lượng sẽ không hề xảy ra. Enzym hoạt động giải trí như chất xúc tác và được cho phép những phản ứng diễn ra với vận tốc nhanh hơn. Enzym cũng được cho phép điều hòa những con đường chuyển hóa nhằm mục đích cung ứng với những biến hóa trong môi trường tự nhiên của tế bào hoặc tín hiệu từ những tế bào khác .
Hệ thống chuyển hóa của một sinh vật cụ thể sẽ xác định chất nào sẽ là chất dinh dưỡng hoặc là chất độc hại với chúng. Ví dụ, một số sinh vật nhân sơ có thể sử dụng hydrogen sulfide như một chất dinh dưỡng, nhưng khí này lại gây độc đối với động vật.[1] Tốc độ chuyển hóa sẽ ảnh hưởng đến lượng thức ăn mà sinh vật yêu cầu, và cũng ảnh hưởng đến cách thức chúng có thể hấp thụ thức ăn đó.
Bạn đang đọc: Trao đổi chất – Wikipedia tiếng Việt
 Adenosine triphosphat (ATP), phân tử được coi là đồng tiền năng lượng của tế bào
Adenosine triphosphat (ATP), phân tử được coi là đồng tiền năng lượng của tế bào
Một đặc điểm nổi bật của quá trình trao đổi chất là sự giống nhau của các con đường và thành phần chuyển hóa cơ bản giữa các loài khác nhau.[2] Ví dụ, tập hợp các axit carboxylic, được biết đến như là sản phẩm trung gian trong chu trình axit citric, có mặt trong tất cả các sinh vật đã biết, được tìm thấy từ các loài chỉ như vi khuẩn đơn bào Escherichia coli đến tận các sinh vật đa bào lớn như voi.[3] Những điểm tương đồng nổi bật trong các con đường trao đổi chất có thể là do sự xuất hiện sớm của chúng trong lịch sử tiến hóa và vẫn được giữ lại vì mang hiệu quả cao.[4][5]
Mục lục
Thành phần hóa sinh chủ chốt[sửa|sửa mã nguồn]
Hầu hết những cấu trúc và thành phần làm ra động vật hoang dã, thực vật hay vi sinh vật được cấu thành từ bốn đại phân tử cơ bản : amino acid, axit nucleic, carbohydrate và lipid ( thường được gọi là chất béo ). Vì những phân tử này rất quan trọng cho sự sống, nên những phản ứng trao đổi chất tập trung chuyên sâu vào việc tạo ra những phân tử này trong quy trình kiến thiết xây dựng tế bào và mô, hoặc phân giải chúng và sử dụng chúng làm nguồn năng lượng qua quy trình tiêu hóa. Các chất hóa sinh này hoàn toàn có thể được phối hợp với nhau để tạo ra những polymerr như DNA và protein, những đại phân tử thiết yếu của sự sống .
Amino acid và protein[sửa|sửa mã nguồn]
 Cấu trúc của một triglycyrideProtein được tạo thành từ chuỗi những amino acid được nối với nhau bởi những link peptide. Nhiều protein là những enzyme tham gia xúc tác những phản ứng hóa học trong quy trình trao đổi chất. Một số protein khác lại có tính năng cấu trúc hoặc công dụng cơ học, ví dụ điển hình như những protein hình thành khung xương tế bào – mạng lưới hệ thống ” giàn giáo ” giúp duy trì hình dạng cả tế bào. [ 6 ] Protein cũng rất quan trọng cho 1 số ít tính năng khác như tín hiệu tế bào liên lạc, cung ứng miễn dịch, bám dính tế bào, luân chuyển dữ thế chủ động qua màng sinh chất và chu kỳ luân hồi tế bào. [ 7 ] amino acid cũng góp thêm phần cho chuyển hóa năng lượng tế bào bằng cách cung ứng nguồn carbon để đi vào quy trình axit citric ( quy trình axit tricarboxylic ), [ 8 ] đặc biệt quan trọng khi nguồn năng lượng chính, ví dụ điển hình như glucose, bị hết sạch hoặc khi những tế bào đang trải qua những stress về chuyển hóa. [ 9 ]
Cấu trúc của một triglycyrideProtein được tạo thành từ chuỗi những amino acid được nối với nhau bởi những link peptide. Nhiều protein là những enzyme tham gia xúc tác những phản ứng hóa học trong quy trình trao đổi chất. Một số protein khác lại có tính năng cấu trúc hoặc công dụng cơ học, ví dụ điển hình như những protein hình thành khung xương tế bào – mạng lưới hệ thống ” giàn giáo ” giúp duy trì hình dạng cả tế bào. [ 6 ] Protein cũng rất quan trọng cho 1 số ít tính năng khác như tín hiệu tế bào liên lạc, cung ứng miễn dịch, bám dính tế bào, luân chuyển dữ thế chủ động qua màng sinh chất và chu kỳ luân hồi tế bào. [ 7 ] amino acid cũng góp thêm phần cho chuyển hóa năng lượng tế bào bằng cách cung ứng nguồn carbon để đi vào quy trình axit citric ( quy trình axit tricarboxylic ), [ 8 ] đặc biệt quan trọng khi nguồn năng lượng chính, ví dụ điển hình như glucose, bị hết sạch hoặc khi những tế bào đang trải qua những stress về chuyển hóa. [ 9 ]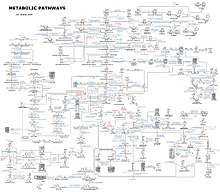 Biểu đồ cho ta thấy một số lượng lớn những con đường chuyển hóa
Biểu đồ cho ta thấy một số lượng lớn những con đường chuyển hóa
Lipid là nhóm chất sinh hóa phong phú nhất. Chức năng cấu trúc chính của chúng là giúp tạo nên những phần của màng sinh học cả bên trong và bên ngoài, ví dụ điển hình như màng tế bào hoặc chúng cũng hoàn toàn có thể dùng làm nguồn năng lượng cho tế bào. [ 7 ] Lipid thường được định nghĩa là những phân tử sinh học kỵ nước hoặc lưỡng phần nhưng lại hoàn toàn có thể tan trong những dung môi hữu cơ như benzene hoặc chloroform. [ 10 ] Chất béo là một nhóm lớn những hợp chất có chứa những axit béo và glycerol. Triacylglyceride là một phân tử được cấu trúc từ một glycerol gắn với ba este axit béo. [ 11 ] Ngoài cấu trúc cơ bản này thì trong tế bào còn sống sót một số ít biến thể, ví dụ điển hình như sphingolipid với mạch khung được thay bằng sphingosine, phospholipid với một trong ba axit béo được thay bằng nhóm ưa nước phosphat. Các steroid như cholesterol cũng là một nhóm lớn khác của lipid. [ 12 ]
 Glucose có thể tồn tại ở cả dạng thẳng và vòng.
Glucose có thể tồn tại ở cả dạng thẳng và vòng.
Carbohydrate hoàn toàn có thể là aldehyde hoặc ketone, với nhiều nhóm hydroxyl được gắn vào, và hoàn toàn có thể sống sót dưới dạng thẳng hoặc vòng. Carbohydrate là nhóm những phân tử sinh học nhiều mẫu mã nhất, và tương thích với nhiều vai trò, ví dụ điển hình như tàng trữ và luân chuyển năng lượng ( tinh bột, glycogen ) hay đóng vai trò là những thành phần cấu trúc ( cellulose ở thực vật, chitin ở động vật hoang dã ). [ 7 ] Các đơn vị chức năng carbohydrate cơ bản được gọi là monosaccharide ( đường đơn ), hoàn toàn có thể kể đến như galactose, fructose, và quan trọng nhất là glucose. Monosaccharide hoàn toàn có thể được link với nhau để tạo thành những polysaccharide ( đường đa ) theo vô số cách khác nhau. [ 13 ]
Hai axit nucleic, DNA và RNA, là những polyme của nucleotide. Mỗi nucleotide gồm một nhóm phosphat gắn với một đường ribose hoặc deoxyribose cùng với một base nitơ. Axit nucleic rất quan trọng cho việc tàng trữ và truyền đạt thông tin di truyền, thông tin di truyền này sẽ được ” diễn giải ” qua quy trình phiên mã và sinh tổng hợp protein. [ 7 ] tin tức này được dữ gìn và bảo vệ bởi những chính sách sửa chữa thay thế DNA và được nhân lên trải qua quy trình sao chép DNA. Nhiều virus lại sử dụng bộ gen RNA, ví dụ điển hình như HIV, và hoàn toàn có thể phiên mã ngược để tạo ra DNA từ bộ gen RNA của virus. [ 14 ] RNA trong ribozyme như thể cắt nối ( spliceosome ) và ribôxôm cũng có hoạt động giải trí tựa như như enzyme vì nó hoàn toàn có thể xúc tác cho những phản ứng hóa học. Các nucleoside riêng không liên quan gì đến nhau được tạo ra bằng cách gắn một nucleobase với đường ribose. Các base này là những hợp chất dị vòng có chứa nitơ, được chia làm hai nhóm là purine hoặc pyrimidine. Nucleotide cũng hoàn toàn có thể hoạt động giải trí như những coenzyme trong phản ứng chuyển-nhóm-chuyển hóa. [ 15 ]
Bài cụ thể : Coenzyme Cấu trúc của coenzyme acetyl-CoA. Nhóm acetyl hoàn toàn có thể chuyển được link với nguyên tử lưu huỳnh ở tận cùng bên trái .Trao đổi chất tương quan đến một lượng lớn những phản ứng hóa học, nhưng hầu hết hoàn toàn có thể được xếp vào một vài loại phản ứng cơ bản tương quan đến việc chuyển những nhóm chức của nguyên tử và link của chúng giữa những phân tử. [ 16 ] Các phản ứng hóa học thường thì này được cho phép những tế bào sử dụng một nhóm nhỏ những chất chuyển hóa trung gian để mang những nhóm chức giữa những phản ứng khác nhau. [ 15 ] Những chất chuyển nhóm trung gian này được gọi là coenzyme. Mỗi loại phản ứng chuyển nhóm này được thực thi bởi một coenzyme đặc hiệu, là cơ chất cho một tập hợp những enzyme tạo ra, và một tập hợp enzyme khác sử dụng chúng. Do đó, những coenzyme này liên tục được tạo ra, sử dụng và sau đó lại được tái tạo. [ 17 ]Một coenzym quan trọng là adenosine triphosphate ( ATP ), ” đồng xu tiền năng lượng ” chung cho tế bào. Nucleotide này được sử dụng để chuyển năng lượng hóa học giữa những phản ứng hóa học khác nhau. Chỉ có một lượng nhỏ ATP trong những tế bào, nhưng chúng được tái tạo liên tục ; mỗi ngày khung hình con người hoàn toàn có thể sử dụng một lượng ATP bằng với khối lượng của mình. [ 17 ] ATP hoạt động giải trí như một cầu nối giữa hai quy trình là dị hóa và đồng nhất. Dị hóa thì hủy hoại những phân tử, còn đồng nhất lại xây nên những phân tử này. Phản ứng dị hóa tạo ra ATP, và phản ứng đồng điệu lại sử dụng ATP này. ATP cũng hoàn toàn có thể đóng vai trò như chất mang nhóm phosphate trong những phản ứng phosphoryl hóa .Vitamin là một loại hợp chất hữu cơ thiết yếu với lượng nhỏ mà không hề tự tổng hợp trong những tế bào. Trong dinh dưỡng ở người, hầu hết những vitamin hoạt động giải trí như coenzyme sau khi sửa đổi ; ví dụ, tổng thể những vitamin tan trong nước được phosphoryl hóa hoặc được tích hợp với nucleotide khi chúng được sử dụng trong tế bào. [ 18 ] Nicotinamide adenine dinucleotide ( NAD + ), một dẫn xuất của vitamin B3 ( niacin ), là một coenzyme quan trọng đóng vai trò làm chất nhận hydro. Có hàng trăm loại enzyme dehydrogenase riêng không liên quan gì đến nhau cho việc vô hiệu những electron khỏi cơ chất của chúng và khử NAD + thành NADH. NADH này lại hoàn toàn có thể sử dụng để khử những cơ chất khác với hoạt động giải trí của enzyme reductase. [ 19 ] Nicotinamide adenine dinucleotide sống sót ở hai dạng ” thân mật ” trong tế bào là NADH và NADPH. Dạng NAD + / NADH quan trọng hơn trong những phản ứng dị hóa, còn dạng NADP + / NADPH được sử dụng trong những phản ứng đồng điệu .
Cấu trúc của coenzyme acetyl-CoA. Nhóm acetyl hoàn toàn có thể chuyển được link với nguyên tử lưu huỳnh ở tận cùng bên trái .Trao đổi chất tương quan đến một lượng lớn những phản ứng hóa học, nhưng hầu hết hoàn toàn có thể được xếp vào một vài loại phản ứng cơ bản tương quan đến việc chuyển những nhóm chức của nguyên tử và link của chúng giữa những phân tử. [ 16 ] Các phản ứng hóa học thường thì này được cho phép những tế bào sử dụng một nhóm nhỏ những chất chuyển hóa trung gian để mang những nhóm chức giữa những phản ứng khác nhau. [ 15 ] Những chất chuyển nhóm trung gian này được gọi là coenzyme. Mỗi loại phản ứng chuyển nhóm này được thực thi bởi một coenzyme đặc hiệu, là cơ chất cho một tập hợp những enzyme tạo ra, và một tập hợp enzyme khác sử dụng chúng. Do đó, những coenzyme này liên tục được tạo ra, sử dụng và sau đó lại được tái tạo. [ 17 ]Một coenzym quan trọng là adenosine triphosphate ( ATP ), ” đồng xu tiền năng lượng ” chung cho tế bào. Nucleotide này được sử dụng để chuyển năng lượng hóa học giữa những phản ứng hóa học khác nhau. Chỉ có một lượng nhỏ ATP trong những tế bào, nhưng chúng được tái tạo liên tục ; mỗi ngày khung hình con người hoàn toàn có thể sử dụng một lượng ATP bằng với khối lượng của mình. [ 17 ] ATP hoạt động giải trí như một cầu nối giữa hai quy trình là dị hóa và đồng nhất. Dị hóa thì hủy hoại những phân tử, còn đồng nhất lại xây nên những phân tử này. Phản ứng dị hóa tạo ra ATP, và phản ứng đồng điệu lại sử dụng ATP này. ATP cũng hoàn toàn có thể đóng vai trò như chất mang nhóm phosphate trong những phản ứng phosphoryl hóa .Vitamin là một loại hợp chất hữu cơ thiết yếu với lượng nhỏ mà không hề tự tổng hợp trong những tế bào. Trong dinh dưỡng ở người, hầu hết những vitamin hoạt động giải trí như coenzyme sau khi sửa đổi ; ví dụ, tổng thể những vitamin tan trong nước được phosphoryl hóa hoặc được tích hợp với nucleotide khi chúng được sử dụng trong tế bào. [ 18 ] Nicotinamide adenine dinucleotide ( NAD + ), một dẫn xuất của vitamin B3 ( niacin ), là một coenzyme quan trọng đóng vai trò làm chất nhận hydro. Có hàng trăm loại enzyme dehydrogenase riêng không liên quan gì đến nhau cho việc vô hiệu những electron khỏi cơ chất của chúng và khử NAD + thành NADH. NADH này lại hoàn toàn có thể sử dụng để khử những cơ chất khác với hoạt động giải trí của enzyme reductase. [ 19 ] Nicotinamide adenine dinucleotide sống sót ở hai dạng ” thân mật ” trong tế bào là NADH và NADPH. Dạng NAD + / NADH quan trọng hơn trong những phản ứng dị hóa, còn dạng NADP + / NADPH được sử dụng trong những phản ứng đồng điệu .
Chất khoáng và cofactor[sửa|sửa mã nguồn]
 Cấu trúc của hemoglobin. Các tiểu đơn vị chức năng protein được tô màu đỏ và xanh dương, và những nhóm heme chứa sắt thì có màu xanh lục. Từ .Các nguyên tố vô cơ cũng đóng vai trò quan trọng trong quy trình trao đổi chất ; 1 số ít thì rất giàu trong tế bào ( ví dụ : natri và kali ) trong khi 1 số ít khác hoạt động giải trí ở nồng độ rất thấp. Khoảng 99 % khối lượng của động vật hoang dã có vú được tạo thành từ những nguyên tố carbon, nitơ, calci, natri, clo, kali, hydro, phospho, oxy và lưu huỳnh. [ 20 ] Các hợp chất hữu cơ ( protein, lipid và carbohydrate ) có hầu hết thành phần là carbon và nitơ ; hầu hết oxy và hydro có mặt dưới dạng nước. [ 20 ]Các nguyên tố vô cơ đa dạng chủng loại đóng vai trò như những ion điện ly. Các ion quan trọng nhất là natri, kali, calci, magiê, chloride, phosphat và ion bicacbonat hữu cơ. Việc duy trì gradient ion đúng mực trên màng tế bào giúp duy trì không thay đổi áp suất thẩm thấu và pH. [ 21 ] Các ion cũng đặc biệt quan trọng quan trọng so với công dụng của tế bào thần kinh và cơ, vì điện thế hoạt động giải trí trong những mô này được tạo ra bằng cách trao đổi những chất điện giải giữa dịch ngoại bào và phần lỏng của tế bào, còn gọi là bào tương. [ 22 ] Các chất điện giải đi vào và rời những tế bào qua những protein trên màng tế bào được gọi là những kênh ion. Ví dụ, hoạt động giải trí co cơ phụ thuộc vào sự di dời của những ion calci, natri và kali nhờ những kênh trên màng sinh chất và những ống T. [ 23 ]Kim loại chuyển tiếp thường xuất hiện với vai trò là những nguyên tố vi lượng trong những sinh vật, kẽm và sắt là những nguyên tố nhiều mẫu mã nhất trong nhóm này. [ 24 ] [ 25 ] Những sắt kẽm kim loại này được sử dụng trong một số ít protein như cofactor và rất thiết yếu cho hoạt động giải trí của những enzyme như catalase và những protein luân chuyển oxy như hemoglobin. [ 26 ] Cofactor sắt kẽm kim loại được gắn chặt vào những vị trí đặc hiệu trong protein ; và mặc dầu cofactor của enzyme hoàn toàn có thể được đổi khác trong quy trình xúc tác, chúng luôn trở về trạng thái khởi đầu vào cuối phản ứng. Các sắt kẽm kim loại vi lượng này được hấp thụ vào sinh vật qua những chất luân chuyển đặc hiệu và nếu chúng chưa được sử dụng : chúng sẽ link với những protein dự trữ như ferritin hoặc metallothionein. [ 27 ] [ 28 ]
Cấu trúc của hemoglobin. Các tiểu đơn vị chức năng protein được tô màu đỏ và xanh dương, và những nhóm heme chứa sắt thì có màu xanh lục. Từ .Các nguyên tố vô cơ cũng đóng vai trò quan trọng trong quy trình trao đổi chất ; 1 số ít thì rất giàu trong tế bào ( ví dụ : natri và kali ) trong khi 1 số ít khác hoạt động giải trí ở nồng độ rất thấp. Khoảng 99 % khối lượng của động vật hoang dã có vú được tạo thành từ những nguyên tố carbon, nitơ, calci, natri, clo, kali, hydro, phospho, oxy và lưu huỳnh. [ 20 ] Các hợp chất hữu cơ ( protein, lipid và carbohydrate ) có hầu hết thành phần là carbon và nitơ ; hầu hết oxy và hydro có mặt dưới dạng nước. [ 20 ]Các nguyên tố vô cơ đa dạng chủng loại đóng vai trò như những ion điện ly. Các ion quan trọng nhất là natri, kali, calci, magiê, chloride, phosphat và ion bicacbonat hữu cơ. Việc duy trì gradient ion đúng mực trên màng tế bào giúp duy trì không thay đổi áp suất thẩm thấu và pH. [ 21 ] Các ion cũng đặc biệt quan trọng quan trọng so với công dụng của tế bào thần kinh và cơ, vì điện thế hoạt động giải trí trong những mô này được tạo ra bằng cách trao đổi những chất điện giải giữa dịch ngoại bào và phần lỏng của tế bào, còn gọi là bào tương. [ 22 ] Các chất điện giải đi vào và rời những tế bào qua những protein trên màng tế bào được gọi là những kênh ion. Ví dụ, hoạt động giải trí co cơ phụ thuộc vào sự di dời của những ion calci, natri và kali nhờ những kênh trên màng sinh chất và những ống T. [ 23 ]Kim loại chuyển tiếp thường xuất hiện với vai trò là những nguyên tố vi lượng trong những sinh vật, kẽm và sắt là những nguyên tố nhiều mẫu mã nhất trong nhóm này. [ 24 ] [ 25 ] Những sắt kẽm kim loại này được sử dụng trong một số ít protein như cofactor và rất thiết yếu cho hoạt động giải trí của những enzyme như catalase và những protein luân chuyển oxy như hemoglobin. [ 26 ] Cofactor sắt kẽm kim loại được gắn chặt vào những vị trí đặc hiệu trong protein ; và mặc dầu cofactor của enzyme hoàn toàn có thể được đổi khác trong quy trình xúc tác, chúng luôn trở về trạng thái khởi đầu vào cuối phản ứng. Các sắt kẽm kim loại vi lượng này được hấp thụ vào sinh vật qua những chất luân chuyển đặc hiệu và nếu chúng chưa được sử dụng : chúng sẽ link với những protein dự trữ như ferritin hoặc metallothionein. [ 27 ] [ 28 ]
Quá trình dị hóa[sửa|sửa mã nguồn]
Dị hóa là tập hợp những quy trình chuyển hóa làm ” phân nhỏ ” những đại phân tử. Chúng cũng gồm có cả quy trình phân giải và oxy hóa những chất dinh dưỡng trong thức ăn. Mục đích của những phản ứng dị hóa là cung ứng năng lượng và những nguyên vật liệu thiết yếu cho những phản ứng đồng điệu – xây nên những phân tử phức tạp hơn. Bản chất đúng chuẩn của những phản ứng dị hóa này là khác nhau so với những sinh vật khác nhau. Do vậy, sinh vật hoàn toàn có thể được phân loại dựa trên nguồn năng lượng và carbon của chúng ( hay nhóm dinh dưỡng chính của chúng ), như trong bảng dưới đây. Các phân tử hữu cơ được sử dụng làm nguồn năng lượng bởi những sinh vật hữu cơ dưỡng, trong khi những sinh vật vô cơ dưỡng lại sử dụng cơ chất vô cơ còn những sinh vật quang dưỡng thì thu nhận ánh sáng mặt trời làm năng lượng hóa học. Tuy nhiên, toàn bộ những dạng trao đổi chất khác nhau phụ thuộc vào những phản ứng oxy hóa khử tương quan đến việc chuyển những electron từ những chất cho điện tử như phân tử hữu cơ, nước, amonia, hydrogen sulfide hoặc những ion chứa sắt sang những chất nhận điện tử như oxy, nitrat hoặc sulfat. [ 29 ] Ở động vật hoang dã, những phản ứng này tương quan đến những phân tử hữu cơ phức tạp được bẻ gãy thành những phân tử đơn thuần hơn, như carbon dioxide và nước. Trong những sinh vật quang hợp, ví dụ điển hình như thực vật và vi trùng lam, những phản ứng chuyển điện tử này không giải phóng năng lượng nhưng được sử dụng như một cách để dự trữ năng lượng hấp thụ từ ánh sáng mặt trời. [ 30 ]
Các phản ứng dị hóa thông dụng nhất ở động vật hoang dã hoàn toàn có thể được chia thành ba quá trình chính. Trong quá trình tiên phong, những đại phân tử hữu cơ, ví dụ điển hình như protein, polysaccharide hoặc lipid, bị tiêu hóa thành những phần nhỏ hơn ở bên ngoài tế bào. Tiếp theo, những phân tử nhỏ này được những tế bào hấp thu và chuyển thành những phân tử nhỏ hơn nữa, thường là acetyl coenzyme A ( acetyl-CoA ), kèm theo giải phóng một số ít năng lượng. Cuối cùng, nhóm acetyl trên CoA bị oxy hóa thành nước và carbon dioxide trong quy trình axit citric và chuỗi luân chuyển electron, giải phóng năng lượng được tàng trữ bằng cách khử coenzyme nicotinamide adenine dinucleotide ( NAD + ) thành NADH .
Tế bào không hề hấp thụ được ngay những đại phân tử như tinh bột, cellulose hoặc protein, những phân tử rất lớn này phải chia thành những phần nhỏ hơn trước khi chúng hoàn toàn có thể được sử dụng trong quy trình chuyển hóa tế bào. Một số loại enzyme phổ cập sẽ giúp tiêu hóa những polymer này. Các enzyme tiêu hóa này gồm có protease giúp tiêu hóa protein thành những amino acid, hay như những glycoside hydrolase sẽ tiêu hóa polysaccharide thành những loại đường đơn hay còn gọi là monosaccharide .Vi khuẩn thì đơn thuần là tiết những enzyme tiêu hóa vào môi trường tự nhiên xung quanh, [ 31 ] [ 32 ] còn động vật hoang dã chỉ tiết ra những enzyme này từ những tế bào được chuyên hóa trong đường ống tiêu hóa, hoàn toàn có thể kể đến như dạ dày, tuyến tụy, và tuyến nước bọt. [ 33 ] Các amino acid hoặc đường được tạo ra nhờ những enzyme ngoại bào này sau đó được đưa vào những tế bào bằng những protein luân chuyển tích cực. [ 34 ] [ 35 ]
Năng lượng từ những hợp chất hữu cơ[sửa|sửa mã nguồn]
 carbohydrate, chất béoSơ đồ đơn thuần cho quy trình phân giải những phân tử : proteinDị hóa carbohydrate là phân giải những phân tử carbohydrate thành những đơn vị chức năng nhỏ hơn. Carbohydrate thường đi vào tế bào sau khi chúng được tiêu hóa thành những monosaccharide. [ 36 ] Khi đã ở bên trong tế bào, con đường chính của phân giải là đường phân ( ” tách đường ” ). Các loại đường như glucose và fructose sau khi tham gia đường phân sẽ được tách thành pyruvate và tạo ra một số ít ATP. [ 37 ] Pyruvate là chất trung gian cho một số ít con đường chuyển hóa khác nhau, nhưng hầu hết chúng sẽ được chuyển thành acetyl-CoA trải qua quy trình đường phân hiếu khí ( có oxy ) và đi vào quy trình axit citric. Mặc dù một số ít ATP được tạo ra trong quy trình axit citric, loại sản phẩm quan trọng nhất của quy trình này là NADH, được tạo thành từ NAD + với điện tử từ acetyl-CoA. Quá trình oxy hóa này giải phóng carbon dioxide là mẫu sản phẩm thải. Trong điều kiện kèm theo yếm khí, đường phân lại tạo ra lactate, nhờ enzyme lactate dehydrogenase tái oxy hóa NADH thành NAD + để liên tục sử dụng trong lần đường phân tiếp theo. Ngoài đường phân, glucose hoàn toàn có thể được phân giải theo con đường khác là con đường pentose phosphate, khử coenzyme NADPH và tạo ra những loại đường pentose như ribose, thành phần đường hoàn toàn có thể cấu trúc nên axit nucleic .
carbohydrate, chất béoSơ đồ đơn thuần cho quy trình phân giải những phân tử : proteinDị hóa carbohydrate là phân giải những phân tử carbohydrate thành những đơn vị chức năng nhỏ hơn. Carbohydrate thường đi vào tế bào sau khi chúng được tiêu hóa thành những monosaccharide. [ 36 ] Khi đã ở bên trong tế bào, con đường chính của phân giải là đường phân ( ” tách đường ” ). Các loại đường như glucose và fructose sau khi tham gia đường phân sẽ được tách thành pyruvate và tạo ra một số ít ATP. [ 37 ] Pyruvate là chất trung gian cho một số ít con đường chuyển hóa khác nhau, nhưng hầu hết chúng sẽ được chuyển thành acetyl-CoA trải qua quy trình đường phân hiếu khí ( có oxy ) và đi vào quy trình axit citric. Mặc dù một số ít ATP được tạo ra trong quy trình axit citric, loại sản phẩm quan trọng nhất của quy trình này là NADH, được tạo thành từ NAD + với điện tử từ acetyl-CoA. Quá trình oxy hóa này giải phóng carbon dioxide là mẫu sản phẩm thải. Trong điều kiện kèm theo yếm khí, đường phân lại tạo ra lactate, nhờ enzyme lactate dehydrogenase tái oxy hóa NADH thành NAD + để liên tục sử dụng trong lần đường phân tiếp theo. Ngoài đường phân, glucose hoàn toàn có thể được phân giải theo con đường khác là con đường pentose phosphate, khử coenzyme NADPH và tạo ra những loại đường pentose như ribose, thành phần đường hoàn toàn có thể cấu trúc nên axit nucleic .
Chất béo được dị hóa qua phản ứng thủy phân thành axit béo tự do và glycerol. Glycerol sẽ đi vào đường phân và các axit béo được phân nhỏ bởi quá trình beta oxy hóa để giải phóng acetyl-CoA, rồi chất này lại đi vào chu trình axit citric. Axit béo sẽ tạo ra nhiều năng lượng hơn so với carbohydrate vì carbohydrate chứa nhiều oxy hơn trong cấu trúc của chúng. Steroid cũng bị phá vỡ bởi một số vi khuẩn bằng quá trình tương tự như quá trình beta oxy hóa, và quá trình phân giải này liên quan đến việc giải phóng một lượng đáng kể acetyl-CoA, propionyl-CoA và pyruvate, tất cả đều có thể được sử dụng bởi tế bào để tạo ra năng lượng. Loài M. tuberculosis (“vi khuẩn lao”) cũng có thể sinh trưởng chỉ với lipid cholesterol là nguồn carbon duy nhất; các gen liên quan đến con đường sử dụng cholesterol được xác định là tối quan trọng trong các giai đoạn lây nhiễm của vi khuẩn này.[38]
amino acid được sử dụng để tổng hợp protein và các phân tử sinh học khác, hoặc cũng có thể bị oxy hóa thành urê và carbon dioxide để sinh năng lượng.[39] Quá trình oxy hóa bắt đầu với việc loại bỏ các nhóm amin bởi một enzyme transaminase. Nhóm amin sẽ được đưa vào chu trình urê, để lại một bộ khung cacbon dưới dạng axit keto. Một số axit keto là chất trung gian trong chu trình axit citric, ví dụ như bước khử amin glutamate để tạo α-ketoglutarate.[40] Các amino acid tạo đường cũng có thể được chuyển đổi thành glucose thông qua con đường tân tạo đường (gluconeogenesis) (thảo luận dưới đây).[41]
Biến đổi năng lượng[sửa|sửa mã nguồn]
Phosphoryl hóa oxy hóa[sửa|sửa mã nguồn]
Trong quy trình phosphoryl hóa oxy hóa, những electron bị tách khỏi những phân tử hữu cơ trong những quy trình như quy trình axit citric sẽ được chuyển tới oxy và giải phóng năng lượng. Năng lượng này sẽ được sử dụng để tổng hợp ATP. Ở sinh vật nhân chuẩn, phosphoryl hóa oxy hóa được triển khai bởi một loạt những protein trên màng ti thể gọi là chuỗi luân chuyển điện tử. Ở sinh vật nhân sơ, những protein tham gia lại được tìm thấy ở màng trong của tế bào. [ 42 ] Các protein này sử dụng năng lượng giải phóng từ việc truyền electron từ những phân tử bị khử như NADH đến oxy để bơm proton qua màng. [ 43 ]
 Cơ chế của ATP synthase. ATP được bộc lộ bằng màu đỏ, ADP và phosphat là màu hồng và tiểu đơn vị chức năng xoay là màu đen .Việc bơm proton ra khỏi chất nền ti thể tạo ra chênh lệch nồng độ proton trên màng tế bào và hình thành một gradient điện hóa. [ 44 ] Theo đúng nguyên tắc khuếch tán, proton sẽ vào lại vào chất nền ty thể ( do nồng độ proton ở xoang gian màng cao hơn trong chất nền ) và đi qua một enzyme gọi là ATP synthase. Dòng proton sẽ khiến một tiểu đơn vị chức năng của enzyme này quay, làm đổi khác hình dạng vị trí hoạt động giải trí của miền synthase và phosphoryl hóa adenosine diphosphate ( ADP ) để tạo thành ATP. [ 17 ]
Cơ chế của ATP synthase. ATP được bộc lộ bằng màu đỏ, ADP và phosphat là màu hồng và tiểu đơn vị chức năng xoay là màu đen .Việc bơm proton ra khỏi chất nền ti thể tạo ra chênh lệch nồng độ proton trên màng tế bào và hình thành một gradient điện hóa. [ 44 ] Theo đúng nguyên tắc khuếch tán, proton sẽ vào lại vào chất nền ty thể ( do nồng độ proton ở xoang gian màng cao hơn trong chất nền ) và đi qua một enzyme gọi là ATP synthase. Dòng proton sẽ khiến một tiểu đơn vị chức năng của enzyme này quay, làm đổi khác hình dạng vị trí hoạt động giải trí của miền synthase và phosphoryl hóa adenosine diphosphate ( ADP ) để tạo thành ATP. [ 17 ]
Năng lượng từ những hợp chất vô cơ[sửa|sửa mã nguồn]
Hóa vô cơ dưỡng là một hình thức chuyển hóa được tìm thấy ở những sinh vật nhân sơ. Khác với quy trình trên, năng lượng thu được từ quy trình oxy hóa là từ những hợp chất vô cơ. Những sinh vật này hoàn toàn có thể sử dụng hydro, [ 45 ] những hợp chất bị khử của lưu huỳnh ( như sulfide, hydrogen sulfide và thiosulfat ), [ 1 ] sắt ( II ) oxit [ 46 ] hoặc amonia [ 47 ] làm nguồn năng lượng khử và chúng sẽ oxy hóa những hợp chất này với những chất nhận electron như oxy hoặc nitrit để tạo năng lượng. [ 48 ] Các quy trình vi sinh này rất quan trọng trong những quy trình sinh địa hóa toàn thế giới như quy trình tạo acetic, nitrat hóa và khử nitơ cũng như rất quan trọng so với độ phì nhiêu của đất. [ 49 ] [ 50 ]
Năng lượng từ ánh sáng mặt trời[sửa|sửa mã nguồn]
Năng lượng trong ánh sáng mặt trời bị ” bẫy ” hay bắt giữ bởi thực vật, vi trùng lam, vi trùng tía, vi trùng lưu huỳnh màu lục và một số ít sinh vật nguyên sinh. Quá trình này thường được tích hợp với việc quy đổi cacbon dioxide thành những hợp chất hữu cơ, như thể một phần của quy trình quang hợp ( sẽ được luận bàn dưới đây ). Tuy nhiên, mạng lưới hệ thống ” bẫy ” năng lượng và mạng lưới hệ thống cố định và thắt chặt cacbon hoàn toàn có thể hoạt động giải trí độc lập trong những sinh vật nhân sơ, ví dụ điển hình như ở vi trùng tía và vi trùng lưu huỳnh màu lục hoàn toàn có thể sử dụng ánh sáng mặt trời làm nguồn năng lượng, trong khi quy đổi giữa cố định và thắt chặt cacbon và lên men những hợp chất hữu cơ. [ 51 ] [ 52 ]Trong nhiều sinh vật, việc ” bẫy ” năng lượng mặt trời có nguyên tắc khá giống với quy trình phosphoryl hóa oxy hóa, vì đều tương quan đến việc tàng trữ năng lượng dưới dạng gradient điện hóa của proton. Lực đẩy proton này sau đó sẽ thôi thúc tổng hợp ATP. [ 17 ] Các electron thiết yếu để quản lý và vận hành chuỗi luân chuyển electron này đến từ những protein-thu-nhận-ánh-sáng được gọi là những TT phản ứng quang hợp hoặc những rhodopsin. Các TT phản ứng được chia thành hai loại tùy thuộc vào loại sắc tố quang hợp hiện hữu ; mà ở hầu hết những vi trùng quang hợp thì chỉ có một loại, trong khi thực vật và vi trùng lam có đến hai loại. [ 53 ]Ở thực vật, tảo và vi trùng lam, mạng lưới hệ thống quang hợp hay quang hệ II sử dụng năng lượng ánh sáng để vô hiệu những electron khỏi nước, giải phóng và thải ra oxy. Các electron sau đó sẽ đi vào phức tạp cytochrome b6f, sử dụng năng lượng của chúng để bơm những proton xuyên qua màng thylakoid trong lục lạp. [ 30 ] Những proton này sẽ vào trở lại qua màng tế bào và đi qua ATP synthase, giống như đã nói ở trên. Các electron sau đó sẽ tới quang hệ I và tiếp đó hoàn toàn có thể được sử dụng để khử coenzyme NADP + thành NADPH để sử dụng trong chu kỳ luân hồi Calvin ( sẽ bàn luận dưới đây ) hoặc được quay vòng để sinh ra thêm ATP. [ 54 ]
Xem thêm thông tin : Đồng hóa
Đồng hóa là tập hợp các quá trình chuyển hóa nhằm “xây dựng”, tổng hợp các phân tử phức tạp với năng lượng được lấy từ các phản ứng dị hóa. Nhìn chung, các phân tử phức tạp tạo thành cấu trúc tế bào được xây dựng dần dần từ các tiền chất nhỏ và đơn giản hơn. Quá trình đồng hóa liên quan đến ba giai đoạn cơ bản. Bước đầu tiên, tổng hợp các tiền chất như amino acid, monosaccharide, isoprenoid và nucleotide, bước thứ hai, hoạt hóa chúng trở thành dạng phản ứng với năng lượng từ ATP, và bước thứ ba, lắp ráp các tiền chất này và tạo nên các phân tử phức tạp như protein, polysaccharides, lipid và axit nucleic.
Các sinh vật khác nhau thì có những cách khác nhau để tổng hợp những chất trong tế bào của mình. Các sinh vật tự dưỡng như thực vật hoàn toàn có thể kiến thiết xây dựng những phân tử hữu cơ phức tạp trong những tế bào như polysaccharide và protein từ những phân tử chỉ đơn thuần như carbon dioxide và nước. Các sinh vật dị dưỡng, mặt khác, để sản xuất những phân tử lớn như vậy lại yên cầu một đầu vào phức tạp hơn, ví dụ điển hình như monosaccharide và amino acid. Các sinh vật hoàn toàn có thể được phân loại hơn nữa dựa trên năng lượng tối ưu cho chúng : sinh vật quang tự dưỡng và quang dị dưỡng thu năng lượng từ ánh sáng mặt trời, trong khi sinh vật hóa tự dưỡng và hóa dị dưỡng lại có được năng lượng từ những phản ứng oxy hóa vô cơ .
Cố định cacbon[sửa|sửa mã nguồn]
 Các tế bào thực vật ( được số lượng giới hạn bởi những vách trên hình ) chứa đầy lục lạp ( xanh lục ), là nơi quang hợpQuang hợp là quy trình tổng hợp cacbohydrat nhờ ánh sáng mặt trời và carbon dioxide ( CO2 ). Ở thực vật, vi trùng lam và tảo, trong quang hợp thải oxy, nước được ” tách ” ra ( gọi là quy trình quang phân li ) và oxy tạo ra như một loại sản phẩm thải. Quá trình này sử dụng ATP và NADPH được tạo ra bởi những TT quang hóa, và như đã miêu tả ở trên, để quy đổi CO2 thành glycerate 3 – phosphate, sau đó chất này hoàn toàn có thể biến hóa thành glucose. Phản ứng cố định và thắt chặt cacbon này được thực thi bởi enzyme RuBisCO và là một phần của quy trình Calvin – Benson. [ 55 ] Có thể tạm nói có ba loại quang hợp xảy ra ở thực vật, cố định và thắt chặt carbon C3, cố định và thắt chặt carbon C4 và quang hợp CAM. Chúng khác nhau theo lộ trình mà CO2 đi vào quy trình Calvin : những cây C3 thì cố định và thắt chặt CO2 trực tiếp, trong khi quang hợp C4 và CAM gắn CO2 vào những hợp chất khác trước, đây là một đặc thù thích nghi để chống chịu với ánh sáng mặt trời nóng bức và điều kiện kèm theo khô hạn. [ 56 ]Trong những sinh vật nhân sơ có quang hợp, những chính sách cho quy trình cố định và thắt chặt cacbon là phong phú hơn. Ở những sinh vật này, cacbon dioxide hoàn toàn có thể được cố định và thắt chặt bởi quy trình Calvin – Benson, quy trình axit citric đảo ngược, [ 57 ] hoặc carboxyl hóa acetyl-CoA. [ 58 ] [ 59 ] Các sinh vật nhân sơ hóa tự dưỡng cũng cố định và thắt chặt CO2 trải qua quy trình Calvin – Benson, nhưng sử dụng năng lượng từ những hợp chất vô cơ để thôi thúc phản ứng. [ 60 ]
Các tế bào thực vật ( được số lượng giới hạn bởi những vách trên hình ) chứa đầy lục lạp ( xanh lục ), là nơi quang hợpQuang hợp là quy trình tổng hợp cacbohydrat nhờ ánh sáng mặt trời và carbon dioxide ( CO2 ). Ở thực vật, vi trùng lam và tảo, trong quang hợp thải oxy, nước được ” tách ” ra ( gọi là quy trình quang phân li ) và oxy tạo ra như một loại sản phẩm thải. Quá trình này sử dụng ATP và NADPH được tạo ra bởi những TT quang hóa, và như đã miêu tả ở trên, để quy đổi CO2 thành glycerate 3 – phosphate, sau đó chất này hoàn toàn có thể biến hóa thành glucose. Phản ứng cố định và thắt chặt cacbon này được thực thi bởi enzyme RuBisCO và là một phần của quy trình Calvin – Benson. [ 55 ] Có thể tạm nói có ba loại quang hợp xảy ra ở thực vật, cố định và thắt chặt carbon C3, cố định và thắt chặt carbon C4 và quang hợp CAM. Chúng khác nhau theo lộ trình mà CO2 đi vào quy trình Calvin : những cây C3 thì cố định và thắt chặt CO2 trực tiếp, trong khi quang hợp C4 và CAM gắn CO2 vào những hợp chất khác trước, đây là một đặc thù thích nghi để chống chịu với ánh sáng mặt trời nóng bức và điều kiện kèm theo khô hạn. [ 56 ]Trong những sinh vật nhân sơ có quang hợp, những chính sách cho quy trình cố định và thắt chặt cacbon là phong phú hơn. Ở những sinh vật này, cacbon dioxide hoàn toàn có thể được cố định và thắt chặt bởi quy trình Calvin – Benson, quy trình axit citric đảo ngược, [ 57 ] hoặc carboxyl hóa acetyl-CoA. [ 58 ] [ 59 ] Các sinh vật nhân sơ hóa tự dưỡng cũng cố định và thắt chặt CO2 trải qua quy trình Calvin – Benson, nhưng sử dụng năng lượng từ những hợp chất vô cơ để thôi thúc phản ứng. [ 60 ]
Cacbohydrat và glycan[sửa|sửa mã nguồn]
Trong quá trình chuyển hóa cacbohydrat, các axit hữu cơ đơn giản có thể được chuyển đổi thành monosaccharide như glucose và sau đó được sử dụng để “lắp ráp” nên các polysaccharide như tinh bột. Quá trình tạo ra glucose từ các hợp chất như pyruvate, lactate, glycerol, glycerate 3-phosphate và amino acid được gọi là tân tạo đường hay gluconeogenesis. Quá trình tân tạo đường biến pyruvate thành glucose-6-phosphate thông qua một loạt các chất trung gian, nhiều chất trong số đó cũng giống với trong đường phân.[37] Tuy nhiên, con đường này không chỉ đơn giản là đảo ngược lại con đường đường phân, vì có một vài bước được xúc tác bởi các enzyme không liên quan gì đến đường phân cả. Điều này là quan trọng vì nó cho phép quá trình hình thành và phân hủy glucose được quy định và điều hòa riêng biệt, và ngăn cản cả hai con đường chạy đồng thời và trở thành một chu trình vô ích (giống như một chiếc xe mà không kiểm soát được “tiến lên” hay “lùi xuống”).[61][62]
Mặc dù chất béo là một cách thông dụng để dự trữ năng lượng, nhưng ở những động vật hoang dã có xương sống thì không hề chuyển hóa lượng chất béo dự trữ này thành glucose trải qua tân tạo đường vì những sinh vật này không hề quy đổi acetyl-CoA thành pyruvate ; thực vật thì hoàn toàn có thể, nhưng động vật hoang dã thì không, chúng thiếu cỗ máy enzym thiết yếu. [ 63 ] Kết quả là, nếu nhịn đói một thời hạn dài, động vật hoang dã có xương sống cần tạo ra những thể xeton từ những axit béo để sửa chữa thay thế glucose vì 1 số ít mô như não không hề chuyển hóa những axit béo. [ 64 ] Ở những dạng sinh vật khác như thực vật và vi trùng, yếu tố chuyển hóa này được xử lý bằng quy trình glyoxylate, đi qua bước decarboxyl hóa trong quy trình axit citric và được cho phép biến hóa acetyl-CoA thành oxaloacetate, chất này hoàn toàn có thể được sử dụng để sản xuất glucose. [ 63 ] [ 65 ]Polysaccharide và glycan được tổng hợp bằng cách bổ trợ tuần tự monosaccharide nhờ enzyme glycosyltransferase. Enzyme này sẽ chuyển đường từ một chất cho đường-phosphate phản ứng như uridine diphosphate glucose ( UDP-glucose ) đến một nhóm nhận hydroxyl trên chuỗi polysaccharide đang được tổng hợp. Vì bất kể nhóm hydroxyl nào trên vòng của cơ chất cũng hoàn toàn có thể là nhóm nhận này, những polysaccharide được tạo ra hoàn toàn có thể có với cấu trúc thẳng hoặc phân nhánh. [ 66 ] Polysaccharide được tạo ra hoàn toàn có thể có những công dụng cấu trúc hoặc trao đổi chất, hoặc được gắn vào những lipid và protein bằng những enzyme gọi là oligosaccharyltransferases. [ 67 ] [ 68 ]
Axit béo, isoprenoid và steroid[sửa|sửa mã nguồn]
Axit béo được tạo ra bởi những enzyme tổng hợp axit béo bằng cách trùng hợp và sau đó là khử đi những đơn vị chức năng acetyl-CoA. Các chuỗi acyl trong những axit béo được lan rộng ra bằng một quy trình phản ứng thêm nhóm acyl, tiên phong là khử để tạo ra rượu, khử nước để tạo thành nhóm alkene và sau đó lại khử tiếp để tạo thành nhóm alkane. Các enzyme sinh tổng hợp axit béo được chia thành hai nhóm : nếu ở động vật hoang dã và nấm, tổng thể những phản ứng tổng hợp axit béo này được thực thi bởi một loại protein loại I đa năng, [ 69 ] thì ở thực vật và vi trùng lại có những enzyme loại II riêng không liên quan gì đến nhau để thực thi từng bước trên con đường. [ 70 ] [ 71 ]Terpene và isoprenoid là một nhóm lớn những chất béo gồm có những carotenoid và tạo thành lớp lớn nhất trong những mẫu sản phẩm tự nhiên đến từ thực vật. [ 72 ] Các hợp chất này được tạo ra bằng cách lắp ráp và cải biến những đơn vị chức năng isoprene được cho từ tiền chất phản ứng isopentenyl pyrophosphate và dimethylallyl pyrophosphate. [ 73 ] Những tiền chất này hoàn toàn có thể được tổng hợp theo nhiều cách khác nhau. Ở động vật hoang dã và cá hồi, con đường mevalonate tạo ra những hợp chất này từ acetyl-CoA, [ 74 ] trong khi ở thực vật và vi trùng, con đường không mevalonate sử dụng pyruvate và glyceraldehyde 3 – phosphate làm cơ chất. [ 73 ] [ 75 ] Một phản ứng quan trọng sử dụng những isoprene được hoạt hóa này là phản ứng sinh tổng hợp steroid. Ở đây, những đơn vị chức năng isoprene được phối hợp với nhau để tạo thành squalene và sau đó được gấp lại và tạo thành một tập hợp những vòng để tạo ra lanosterol. [ 76 ] Lanosterol sau đó hoàn toàn có thể được quy đổi thành những steroid khác như cholesterol và ergosterol. [ 76 ] [ 77 ]
Các sinh vật khác nhau có những điểm khác nhau về khả năng tổng hợp 20 amino acid thông thường. Hầu hết vi khuẩn và thực vật có thể tổng hợp tất cả hai mươi loại này, nhưng động vật có vú chỉ có thể tổng hợp mười một amino acid không thiết yếu, vì vậy mà chín amino acid thiết yếu còn lại phải được lấy từ thực phẩm.[7] Một số loài ký sinh đơn giản, chẳng hạn như vi khuẩn Mycoplasma pneumoniae, không có quá trình tổng hợp amino acid và sẽ lấy amino acid trực tiếp từ vật chủ của chúng.[78] Tất cả các amino acid được tổng hợp từ các chất trung gian trong quá trình đường phân, chu trình axit citric hoặc con đường pentose phosphat. Nitơ được cung cấp bởi glutamate và glutamine. Tổng hợp amino acid phụ thuộc vào sự hình thành của axit alpha-keto thích hợp, sau đó được chuyển thành dạng amino acid.[79]
amino acid được tạo thành protein bằng cách lắp ráp với nhau để tạo thành một chuỗi link peptit. Các protein khác nhau do có những trình tự khác nhau của chuỗi bên amino acid : đây chính là cấu trúc bậc một của protein. Cũng giống như những vần âm của bảng vần âm hoàn toàn có thể được phối hợp để tạo thành một loạt những từ vô tận, những amino acid hoàn toàn có thể được link thành những trình tự khác nhau để tạo thành một lượng rất lớn những protein. Protein được tạo ra từ những amino acid, những amino acid này đã được hoạt hóa bằng cách gắn vào một phân tử tRNA qua một link este. Tiền chất aminoacyl-tRNA này được tạo ra trong một phản ứng cần năng lượng ATP và được thực thi nhờ một aminoacyl tRNA synthetase. [ 80 ] Sau đó, aminoacyl-tRNA này là cơ chất cho ribosome, sẽ giúp tích hợp amino acid vào chuỗi protein đang lê dài, dựa vào RNA thông tin đang được dịch mã. [ 81 ]
Tổng hợp và ” cứu vãn ” nucleotide[sửa|sửa mã nguồn]
Nucleotide được tạo thành từ các amino acid, carbon dioxide và axit formic trong các con đường đòi hỏi phải có một lượng lớn năng lượng chuyển hóa.[82] Do đó, hầu hết các sinh vật đều có hệ thống hiệu quả để “cứu vãn” các nucleotide đã được hình thành trước đó.[82][83] Purine được tổng hợp dưới dạng các nucleoside (các base gắn liền với ribose).[84] Cả adenine và guanine đều được tạo ra từ tiền chất nucleoside inosine monophosphate, được tổng hợp bằng cách sử dụng các nguyên tử từ các amino acid glycine, glutamine và axit aspartic, cũng như formate được chuyển từ coenzyme tetrahydrofolate. Pyrimidine, mặt khác, được tổng hợp từ các thể orotate, được tạo thành từ glutamine và aspartate.[85]
Chất lạ sinh học và chuyển hóa oxy hóa-khử[sửa|sửa mã nguồn]
Tất cả các sinh vật thường xuyên tiếp xúc với các hợp chất mà chúng không thể sử dụng làm chất dinh dưỡng cũng như không có chức năng trao đổi chất và do đó, sẽ thật là có hại nếu các chất này được tích tụ trong các tế bào. Những hợp chất có khả năng gây hại này được gọi là chất lạ sinh học (xenobiotic).[86] Một số ví dụ chẳng hạn như thuốc tổng hợp, chất độc tự nhiên và thuốc kháng sinh, các chất này được giải độc bởi một tập hợp các enzyme chuyên biệt. Ở người, các enzyme này bao gồm các loại cytochrome P450 oxidase,[87] UDP-glucuronosyltransferase,[88] và glutathione S-transferase.[89] Hệ thống enzyme này hoạt động trong ba pha, đầu tiên là oxy hóa các chất lạ này (pha I), sau đó là liên hợp các nhóm giúp hòa tan trong nước lên phân tử (pha II). Chất lạ đã qua xử lý sau đó có thể được bơm ra khỏi tế bào, các sinh vật đa bào có thể chuyển hóa thêm sản phẩm này trước khi được bài tiết ra ngoài (pha III). Trong sinh thái học, các phản ứng này đặc biệt quan trọng trong việc phân hủy sinh học các chất gây ô nhiễm và xử lý sinh học đất bị ô nhiễm cũng như sự cố tràn dầu.[90] Nhiều phản ứng ở vi sinh vật giống với phản ứng ở các sinh vật đa bào, nhưng với sự đa dạng đáng kinh ngạc của thế giới vi sinh, các vi sinh vật có thể xử lý nhiều chất lạ sinh học hơn so với các sinh vật đa bào. Nhờ thế mà vi sinh vật có thể sử dụng để phân hủy các chất ô nhiễm hữu cơ bền vững như các hợp chất clo hữu cơ.[91]
Một yếu tố tương quan so với sinh vật hiếu khí là stress oxy hóa. [ 92 ] Trong trường hợp này, những quy trình như phosphoryl hóa oxy hóa hay hình thành link disulfide trong quy trình cuộn gấp protein tạo ra những gốc oxy phản ứng như hydrogen peroxide. [ 93 ] Các nhóm oxy hóa gây hại này được vô hiệu bởi những chất chống oxy hóa như glutathione và những loại enzyme như catalase và peroxidase. [ 94 ] [ 95 ]
Nhiệt động lực học của sinh vật[sửa|sửa mã nguồn]
Các sinh vật vẫn phải tuân theo những định luật về nhiệt động lực học, tương quan đến quy trình truyền nhiệt và sinh công. Định luật thứ hai của nhiệt động lực học phát biểu rằng : trong bất kể mạng lưới hệ thống kín nào, lượng entropy ( đơn vị chức năng đo ” hỗn loạn ” ) không hề giảm. Thoạt nhìn, mức độ tổ chức triển khai tuyệt vời của sinh vật sống có vẻ như xích míc với định luật này ( vì không ” hỗn loạn ” lắm ), điều này vẫn là hoàn toàn có thể vì tổng thể những sinh vật là những mạng lưới hệ thống mở, có những quy trình trao đổi vật chất và năng lượng với thiên nhiên và môi trường xung quanh. Do đó, những mạng lưới hệ thống sống không ở trạng thái cân đối, nhưng thay vào đó là những mạng lưới hệ thống tiêu tán duy trì mức độ phức tạp cao của sinh vật bằng cách làm tăng entropy lớn hơn trong thiên nhiên và môi trường của chúng. [ 96 ] Quá trình trao đổi chất của tế bào đạt được điều này bằng cách liên kết những quy trình dị hóa xảy ra tự phát với những quy trình đồng nhất không tự phát. Trong những thuật ngữ nhiệt động lực học, quy trình trao đổi chất hay chuyển hóa duy trì ” trật tự ” bằng cách tạo ra ” hỗn loạn “. [ 97 ] Có thể nói, sinh vật là những hòn đảo entropy thấp ( ” trật tự ” ) giữa môi trường tự nhiên có entropy luôn tăng lên ( ” hỗn loạn ” ) .
Điều hòa và trấn áp[sửa|sửa mã nguồn]
Vì môi trường của hầu hết các sinh vật liên tục thay đổi, các phản ứng trao đổi chất phải được điều chỉnh hiệu quả để duy trì một tập hợp các điều kiện liên tục trong các tế bào, trạng thái này gọi là cân bằng nội môi.[98][99] Điều hòa chuyển hóa cũng cho phép các sinh vật phản ứng với các tín hiệu và tương tác tích cực với môi trường sống của chúng.[100] Có hai khái niệm rất quan trọng để hiểu về cách thức các con đường trao đổi chất được kiểm soát, và hai khái niệm này liên kết chặt chẽ với nhau. Thứ nhất, điều hòa hoạt động enzyme trong một con đường chuyển hóa là làm tăng hoặc giảm hoạt tính của chúng để đáp ứng với tín hiệu. Thứ hai, sự kiểm soát được thực hiện bởi enzyme này là hiệu quả mà những thay đổi trong hoạt tính của nó có tác động lên tốc độ tổng thể của con đường (thông lượng qua con đường).[101] Ví dụ, một enzyme có thể có những thay đổi lớn trong hoạt tính (tức là nó được điều hòa ở mức cao) nhưng nếu những thay đổi này ít ảnh hưởng đến thông lượng của một con đường chuyển hóa, thì enzyme này không liên quan nhiều đến việc kiểm soát con đường này.[102]
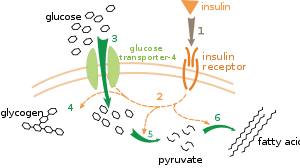 Ảnh hưởng của insulin lên sự hấp thụ và chuyển hóa glucose. Insulin link với thụ thể của chúng ( 1 ), điều này khởi đầu nhiều đợt hoạt hóa protein ( 2 ). Một số phản ứng gồm có : chuyển vị Glut-4 luân chuyển đến màng sinh chất và kênh glucose ( 3 ), tổng hợp glycogen ( 4 ), đường phân ( 5 ) và tổng hợp axit béo ( 6 ) .
Ảnh hưởng của insulin lên sự hấp thụ và chuyển hóa glucose. Insulin link với thụ thể của chúng ( 1 ), điều này khởi đầu nhiều đợt hoạt hóa protein ( 2 ). Một số phản ứng gồm có : chuyển vị Glut-4 luân chuyển đến màng sinh chất và kênh glucose ( 3 ), tổng hợp glycogen ( 4 ), đường phân ( 5 ) và tổng hợp axit béo ( 6 ) .
Có nhiều mức độ điều hòa trao đổi chất. Trong điều hòa nội tại hay bên trong, các con đường trao đổi chất tự điều chỉnh để đáp ứng với những thay đổi về mức độ cơ chất hoặc sản phẩm; ví dụ, nếu số lượng sản phẩm giảm thì có thể tăng thông lượng qua con đường để bù đắp.[101] Kiểu điều hòa này thường liên quan đến điều hòa dị lập thể trong hoạt tính của nhiều enzyme có trong các con đường chuyển hóa này.[103] Điều hòa bên ngoài lại liên quan đến việc tế bào ở một sinh vật đa bào thay đổi mức độ trao đổi chất của nó để đáp ứng với tín hiệu đến từ các tế bào khác. Những tín hiệu này thường ở dạng các chất truyền tin hòa tan như hormone hoặc các yếu tố tăng trưởng và được nhận diện bởi các thụ thể đặc hiệu trên bề mặt tế bào.[104] Những tín hiệu này sau đó được truyền trong tế bào bởi các hệ thống chất truyền tin thứ hai và thường liên quan đến quá trình phosphoryl hóa protein.[105]
Một ví dụ được hiểu rất rõ về trấn áp bên ngoài là điều hòa chuyển hóa glucose trải qua hormone insulin. [ 105 ] Insulin được sản xuất để cung ứng với việc ngày càng tăng nồng độ glucose trong máu ( hay tăng đường huyết ). Các hormone này sẽ gắn với những thụ thể insulin trên màng tế bào và từ đó kích hoạt một loạt những protein kinase làm cho những tế bào hấp thu glucose đồng thời chuyển hóa đường này thành những phân tử dự trữ như axit béo và glycogen. [ 106 ] Quá trình chuyển hóa glycogen được trấn áp bởi hoạt động giải trí của hai enzyme là phosphorylase, một loại enzyme phân giải glycogen, và glycogen synthase, một loại enzyme giúp tạo ra glycogen. Hai enzyme này được kiểm soát và điều chỉnh theo kiểu nghịch đảo : nếu như phosphoryl hóa làm ức chế glycogen synthase, thì lại hoạt hóa phosphorylase. Insulin gây tổng hợp glycogen bằng cách kích hoạt phosphatase protein và làm giảm quy trình phosphoryl hóa những enzyme này. [ 107 ]
Một số con đường chuyển hóa TT mà ta vừa nhắc đến ở trên, ví dụ điển hình như đường phân và quy trình axit citric, là xuất hiện ở cả ba lãnh giới của những sinh vật sống và hiện hữu trong tổ tiên chung phổ cập sau cuối. [ 3 ] [ 108 ] Tế bào tổ tiên này là sinh vật nhân sơ và hoàn toàn có thể là một sinh vật sinh mêtan với một lượng lớn những con đường chuyển hóa amino acid, nucleotide, carbohydrate và lipid. [ 109 ] [ 110 ] Một số con đường cổ xưa vẫn được duy trì đến tận giờ đây. Quá trình tiến hóa hoàn toàn có thể đã tinh lọc những con đường này vì tính tối ưu của chúng trong xử lý những yếu tố chuyển hóa đơn cử, ví dụ điển hình như với đường phân và quy trình axit citric : hai quy trình này tạo ra những mẫu sản phẩm sau cuối với hiệu suất cao cao mà số ” bước ” ( số phản ứng ) là tối thiểu [ 4 ] [ 5 ] Con đường chuyển hóa tiên phong dựa trên enzyme hoàn toàn có thể là một phần trong quy trình trao đổi chất nucleotide purine, còn những con đường chuyển hóa trước đó là một phần của quốc tế RNA cổ đại. [ 111 ]Nhiều quy mô đã được đề xuất kiến nghị để diễn đạt những chính sách mà theo đó những con đường trao đổi chất mới được tăng trưởng. Trong số này hoàn toàn có thể kể đến như : bổ trợ thêm những enzyme mới vào một con đường tổ tiên ngắn, lặp lại và phân kỳ cho hàng loạt con đường, hoặc là tuyển thêm những enzyme đã sống sót và cải biến chúng thành một con đường phản ứng mới. [ 112 ] Tầm quan trọng tương đối của những chính sách này là không rõ ràng, nhưng những điều tra và nghiên cứu gen đã chỉ ra rằng những enzyme trong một con đường hoàn toàn có thể có một tổ tiên chung, cho thấy rằng nhiều con đường đã tăng trưởng từ từ với những công dụng mới được hình thành từ những bước đã sống sót trong con đường trước đó. [ 113 ] Một quy mô thay thế sửa chữa xuất phát từ những điều tra và nghiên cứu theo dõi sự tiến hóa của cấu trúc protein trong những mạng lưới chuyển hóa, quy mô này đã gợi ý rằng : những enzyme được tuyển vào là rất thông dụng, hay tức là ” mượn ” những enzyme để triển khai những tính năng tương tự như trong những con đường trao đổi chất khác nhau ( chứng cứ trong cơ sở tài liệu MANET ) [ 114 ] dẫn đến tiến hóa khảm nhờ enzyme. [ 115 ] Khả năng thứ ba là một số ít phần của quy trình trao đổi chất hoàn toàn có thể sống sót dưới dạng ” module ” hoàn toàn có thể được tái sử dụng trong những con đường khác nhau và triển khai những tính năng tương tự như trên những phân tử khác nhau. [ 115 ]Cũng như tiến hóa của giúp hình thành những con đường trao đổi chất mới, tiến hóa cũng hoàn toàn có thể làm mất một số ít tính năng trao đổi chất. Ví dụ, ở 1 số ít ký sinh trùng, những quy trình chuyển hóa không thiết yếu cho việc sống sót bị mất và những amino acid, nucleotide và carbohydrate hoàn hảo hoàn toàn có thể được lấy khi ” thu dọn ” từ vật chủ. [ 116 ] Khả năng chuyển hóa tối thiểu tựa như cũng được tìm thấy trong những sinh vật nội cộng sinh. [ 117 ]
Nghiên cứu và ứng dụng[sửa|sửa mã nguồn]
 Arabidopsis thaliana. Enzyme và chất chuyển hóa được hiển thị dưới dạng hình vuông màu đỏ và tương tác giữa chúng là đường màu đen.Mạng lưới trao đổi chất của quy trình axit citric của. Enzyme và chất chuyển hóa được hiển thị dưới dạng hình vuông vắn màu đỏ và tương tác giữa chúng là đường màu đen .Theo giải pháp cổ xưa trước đây, quy trình trao đổi chất được nghiên cứu và điều tra theo giải pháp quy giản, tức là tập trung chuyên sâu vào chỉ một con đường chuyển hóa duy nhất. Đặc biệt có giá trị là giải pháp sử dụng những mẫu dò phóng xạ trên hàng loạt khung hình, mô và tế bào, nhằm mục đích xác lập con đường đi từ tiền chất đến mẫu sản phẩm sau cuối bằng cách xác lập những loại sản phẩm trung gian và mẫu sản phẩm sau cuối mang dấu phóng xạ. [ 118 ] Các enzyme xúc tác những phản ứng hóa học này sau đó hoàn toàn có thể được phân lập, động học của những phân tử này cũng như cung ứng của chúng với những chất ức chế đã được nghiên cứu và điều tra. Cách tiếp cận song song là xác lập những phân tử nhỏ trong tế bào hoặc mô ; bộ hoàn hảo những phân tử này được thì gọi là tập chuyển hóa. Nhìn chung, những điều tra và nghiên cứu này đưa ra một cái nhìn tốt về cấu trúc và công dụng của những con đường trao đổi chất đơn thuần, nhưng không đủ khi vận dụng cho những mạng lưới hệ thống phức tạp hơn như sự trao đổi chất của cả một tế bào hoàn hảo. [ 119 ]Để có sáng tạo độc đáo về sự phức tạp của những mạng chuyển hóa trong những tế bào với hàng ngàn enzyme khác nhau, ta hoàn toàn có thể nhìn vào hình biểu lộ tương tác chỉ có 43 protein và 40 chất chuyển hóa ở bên phải, trong khi hệ gen phân phối một list chứa tới 45.000 gen. [ 120 ] Tuy vậy, ở thời gian hiện tại, ta chỉ hoàn toàn có thể sử dụng tài liệu di truyền này để tái tạo lại những mạng hoàn hảo những phản ứng sinh hóa và tạo ra những quy mô toán học toàn diện và tổng thể hơn là hoàn toàn có thể lý giải và Dự kiến hành vi của những tương tác này. [ 121 ] Các quy mô này đặc biệt quan trọng can đảm và mạnh mẽ khi được tích hợp thêm những tài liệu về con đường và chất chuyển hóa thu được trải qua những giải pháp cổ xưa kèm với tài liệu về biểu lộ gen từ những điều tra và nghiên cứu về vi dãy DNA và protein học. [ 122 ] Sử dụng những kỹ thuật này, một quy mô trao đổi chất ở người đã được tạo ra, quy mô này sẽ chỉ hướng cho việc việc mày mò thuốc và nghiên cứu sinh hóa trong tương lai. [ 123 ] Các quy mô này hiện đang được sử dụng trong nghiên cứu và phân tích mạng lưới chuyển hóa, nhằm mục đích phân loại những bệnh ở người thành những nhóm có chung protein hoặc chất chuyển hóa. [ 124 ] [ 125 ]Các mạng trao đổi chất vi trùng là một ví dụ điển hình nổi bật về tổ chức triển khai ” hình nơ “, [ 126 ] [ 127 ] [ 128 ] một mạng lưới hệ thống có năng lực thu nhập vào một lượng những chất dinh dưỡng và sản xuất rất nhiều loại sản phẩm và những đại phân tử phức tạp, nhưng sử dụng rất ít chất trung gian. Do nguồn vào và đầu ra với rất nhiều chất nhưng số lượng chất trung gian ít nên nếu trình diễn trực quan, mạng lưới hệ thống này như bị thắt ở giữa ( giống chiếc nơ ) .Một ứng dụng công nghệ tiên tiến chính của thông tin này là kỹ thuật trao đổi chất. Ở đây, những sinh vật như nấm men, thực vật hoặc vi trùng được biến đổi gen và trở thành những công cụ cực kỳ hữu dụng trong công nghệ sinh học. Chúng hoàn toàn có thể hỗ trợ sản xuất những loại thuốc như kháng sinh hoặc hóa chất công nghiệp như 1,3 – propanediol và axit shikimic. [ 129 ] Những biến hóa di truyền này thường nhằm mục đích mục tiêu giảm lượng năng lượng được sử dụng để sản xuất loại sản phẩm, tăng sản lượng và giảm thiểu chất thải. [ 130 ]
Arabidopsis thaliana. Enzyme và chất chuyển hóa được hiển thị dưới dạng hình vuông màu đỏ và tương tác giữa chúng là đường màu đen.Mạng lưới trao đổi chất của quy trình axit citric của. Enzyme và chất chuyển hóa được hiển thị dưới dạng hình vuông vắn màu đỏ và tương tác giữa chúng là đường màu đen .Theo giải pháp cổ xưa trước đây, quy trình trao đổi chất được nghiên cứu và điều tra theo giải pháp quy giản, tức là tập trung chuyên sâu vào chỉ một con đường chuyển hóa duy nhất. Đặc biệt có giá trị là giải pháp sử dụng những mẫu dò phóng xạ trên hàng loạt khung hình, mô và tế bào, nhằm mục đích xác lập con đường đi từ tiền chất đến mẫu sản phẩm sau cuối bằng cách xác lập những loại sản phẩm trung gian và mẫu sản phẩm sau cuối mang dấu phóng xạ. [ 118 ] Các enzyme xúc tác những phản ứng hóa học này sau đó hoàn toàn có thể được phân lập, động học của những phân tử này cũng như cung ứng của chúng với những chất ức chế đã được nghiên cứu và điều tra. Cách tiếp cận song song là xác lập những phân tử nhỏ trong tế bào hoặc mô ; bộ hoàn hảo những phân tử này được thì gọi là tập chuyển hóa. Nhìn chung, những điều tra và nghiên cứu này đưa ra một cái nhìn tốt về cấu trúc và công dụng của những con đường trao đổi chất đơn thuần, nhưng không đủ khi vận dụng cho những mạng lưới hệ thống phức tạp hơn như sự trao đổi chất của cả một tế bào hoàn hảo. [ 119 ]Để có sáng tạo độc đáo về sự phức tạp của những mạng chuyển hóa trong những tế bào với hàng ngàn enzyme khác nhau, ta hoàn toàn có thể nhìn vào hình biểu lộ tương tác chỉ có 43 protein và 40 chất chuyển hóa ở bên phải, trong khi hệ gen phân phối một list chứa tới 45.000 gen. [ 120 ] Tuy vậy, ở thời gian hiện tại, ta chỉ hoàn toàn có thể sử dụng tài liệu di truyền này để tái tạo lại những mạng hoàn hảo những phản ứng sinh hóa và tạo ra những quy mô toán học toàn diện và tổng thể hơn là hoàn toàn có thể lý giải và Dự kiến hành vi của những tương tác này. [ 121 ] Các quy mô này đặc biệt quan trọng can đảm và mạnh mẽ khi được tích hợp thêm những tài liệu về con đường và chất chuyển hóa thu được trải qua những giải pháp cổ xưa kèm với tài liệu về biểu lộ gen từ những điều tra và nghiên cứu về vi dãy DNA và protein học. [ 122 ] Sử dụng những kỹ thuật này, một quy mô trao đổi chất ở người đã được tạo ra, quy mô này sẽ chỉ hướng cho việc việc mày mò thuốc và nghiên cứu sinh hóa trong tương lai. [ 123 ] Các quy mô này hiện đang được sử dụng trong nghiên cứu và phân tích mạng lưới chuyển hóa, nhằm mục đích phân loại những bệnh ở người thành những nhóm có chung protein hoặc chất chuyển hóa. [ 124 ] [ 125 ]Các mạng trao đổi chất vi trùng là một ví dụ điển hình nổi bật về tổ chức triển khai ” hình nơ “, [ 126 ] [ 127 ] [ 128 ] một mạng lưới hệ thống có năng lực thu nhập vào một lượng những chất dinh dưỡng và sản xuất rất nhiều loại sản phẩm và những đại phân tử phức tạp, nhưng sử dụng rất ít chất trung gian. Do nguồn vào và đầu ra với rất nhiều chất nhưng số lượng chất trung gian ít nên nếu trình diễn trực quan, mạng lưới hệ thống này như bị thắt ở giữa ( giống chiếc nơ ) .Một ứng dụng công nghệ tiên tiến chính của thông tin này là kỹ thuật trao đổi chất. Ở đây, những sinh vật như nấm men, thực vật hoặc vi trùng được biến đổi gen và trở thành những công cụ cực kỳ hữu dụng trong công nghệ sinh học. Chúng hoàn toàn có thể hỗ trợ sản xuất những loại thuốc như kháng sinh hoặc hóa chất công nghiệp như 1,3 – propanediol và axit shikimic. [ 129 ] Những biến hóa di truyền này thường nhằm mục đích mục tiêu giảm lượng năng lượng được sử dụng để sản xuất loại sản phẩm, tăng sản lượng và giảm thiểu chất thải. [ 130 ]
Thuật ngữ metabolism (chuyển hóa) có nguồn gốc từ tiếng Hy Lạp Μεταβολισμός – “Metabolismos” với nghĩa “thay đổi”, hoặc “lật đổ”.[131]
 Mô hình trao đổi chất của Aristole với dạng dòng chảy mở
Mô hình trao đổi chất của Aristole với dạng dòng chảy mở
Trong cuốn Các phần của động vật, Aristotle đã viết đẩy đủ chi tiết từ quan điểm của ông về quá trình trao đổi chất dưới dạng một mô hình dòng chảy mở. Ông tin rằng ở mỗi giai đoạn của quá trình biến đổi, nguyên liệu từ thực phẩm đã được biến đổi, nhiệt được giải phóng thì tượng trưng cho nguyên tố lửa trong cổ điển, còn các phần thừa được bài tiết dưới dạng nước tiểu, mật hoặc phân.[132]
![]() Santorio với chiếc cân khung hình của mình, một bước đi tiên phong trong điều tra và nghiên cứu trao đổi chất ở người .
Santorio với chiếc cân khung hình của mình, một bước đi tiên phong trong điều tra và nghiên cứu trao đổi chất ở người .
Ibn al-Nafis mô tả về trao đổi chất trong tác phẩm năm 1260 của ông với tên là Al-Risalah al-Kamiliyyah fil Siera al-Nabawiyyah (“Lập luận của Kamil cho tiểu sử của vị Tiên tri”), trong đó có những chữ sau đây “Cả cơ thể và các bộ phận của cơ thể đều ở trong trạng thái phá hủy và xây dựng liên tục, nên sự thay đổi tất yếu là vĩnh viễn”[133] Lịch sử nghiên cứu khoa học về quá trình trao đổi chất đã kéo dài vài thế kỷ. Mục tiêu nghiên cứu cũng đã chuyển từ xem xét toàn bộ cơ thể động vật trong các nghiên cứu ban đầu, đến chỉ xét những phản ứng trao đổi chất riêng biệt trong nghiên cứu hóa sinh hiện đại. Các thí nghiệm được kiểm soát đầu tiên về quá trình chuyển hóa của con người được xuất bản bởi Santorio Santorio vào năm 1614 trong cuốn sách Ars de statica medicina.[134] Santorio đã mô tả cách ông tự cân khối lượng chính mình trước và sau khi ăn, ngủ, làm việc, quan hệ tình dục, ăn chay, uống rượu và bài tiết. Nhà y học này đã phát hiện ra rằng hầu hết thức ăn mà ông ăn vào bị mất thông qua cái mà ông gọi là “mồ hôi không thể nhận biết”.
Trong những nghiên cứu và điều tra khởi đầu này, những chính sách của những quy trình trao đổi chất này chưa được xác lập và sống sót học thuyết duy sinh cho rằng : sống sót ” lực sống ” giúp tinh chỉnh và điều khiển những mô sống. [ 135 ] Vào thế kỷ 19, khi điều tra và nghiên cứu quy trình lên men đường thành rượu, Louis Pasteur Tóm lại rằng quy trình lên men được xúc tác bởi những chất trong tế bào nấm men mà ông gọi là ” yếu tố lên men “. Ông viết rằng ” lên men rượu là một quy trình tương quan với sự sống và tổ chức triển khai của những tế bào nấm men, chứ không phải là với cái chết hoặc sự hư hỏng của tế bào. ” [ 136 ] Phát hiện này, cùng với ấn phẩm của Friedrich Wöhler vào năm 1828 cho một bài báo về việc tổng hợp hóa học thành công xuất sắc urê, [ 137 ] đáng chú ý quan tâm vì đây là hợp chất hữu cơ tiên phong được tạo ra từ những tiền chất trọn vẹn vô cơ. Điều này chứng tỏ rằng những hợp chất hữu cơ và những phản ứng hóa học được tìm thấy trong những tế bào sống không độc lạ về mặt nguyên tắc so với bất kể góc nhìn nào khác của hóa học .Việc mày mò ra những enzyme vào đầu thế kỷ 20 bởi Eduard Buchner đã tách việc nghiên cứu và điều tra phản ứng hóa học của trao đổi chất ra khỏi việc điều tra và nghiên cứu sinh học tế bào, và ghi lại sự khởi đầu của bộ môn sinh hóa. [ 138 ] Khối lượng kiến thức sinh hóa đã tăng trưởng nhanh gọn trong suốt đầu thế kỷ 20. Một trong những nhà sinh hóa tân tiến có góp sức nhất là Hans Krebs, ông đã góp phần rất lớn cho điều tra và nghiên cứu về chuyển hóa. [ 139 ] Ông phát hiện ra quy trình urê và sau đó, khi thao tác với Hans Kornberg, ông tìm ra quy trình axit citric và quy trình glyoxylate. [ 65 ] [ 140 ] Nghiên cứu sinh hóa tân tiến đã được tương hỗ rất nhiều bởi sự tăng trưởng của những kỹ thuật mới như sắc ký, nhiễu xạ tia X, quang phổ NMR, ghi lại phóng xạ, kính hiển vi điện tử và mô phỏng động lực phân tử. Những kỹ thuật này đã được cho phép phát hiện và nghiên cứu và phân tích chi tiết cụ thể về nhiều phân tử và con đường chuyển hóa trong tế bào .
Dẫn nhập
- Rose, S. và Mileusnic, R ., The Chemistry of Life. (Hóa học sự sống.) (Penguin Press Science, 1999), ISBN 0-14-027273-9
- Schneider, E. D. và Sagan, D ., Into the Cool: Energy Flow, Thermodynamics, and Life. (Tiến vào vùng lạnh: Dòng năng lượng, nhiệt động lực học, và sự sống.) (Nhà xuất bản Đại học Chicago, 2005), ISBN 0-226-73936-8
- Lane, N ., Oxygen: The Molecule that Made the World. (Oxygen: Phân tử cấu thành thế giới.) (Nhà xuất bản Đại học Oxford, Hoa Kỳ, 2004), ISBN 0-19-860783-0
Nâng cao
- Price, N. và Stevens, L ., Fundamentals of Enzymology: Cell and Molecular Biology of Catalytic Proteins. (Cơ sở về Enzyme học: Sinh học cấp độ tế bào và phân tử của các chất xúc tác protein.) (Nhà xuất bản Đại học Oxford, 1999), ISBN 0-19-850229-X
- Berg, J. Tymoczko, J. và Stryer, L ., Biochemistry. (Hóa sinh.) (W. H. Freeman and Company, 2002), ISBN 0-7167-4955-6
- Cox, M. và Nelson, D. L ., Lehninger Principles of Biochemistry. (Nguyên lý Lehninger của hóa sinh.) (Palgrave Macmillan, 2004), ISBN 0-7167-4339-6
- Brock, T. D.Madigan, M. T. Martinko, J. và Parker J ., Brock’s Biology of Microorganisms. (Sinh học của Brock về vi sinh vật.) (Benjamin Cummings, 2002), ISBN 0-13-066271-2
- Da Silva, J.J.R.F. và Williams, R. J. P ., The Biological Chemistry of the Elements: The Inorganic Chemistry of Life. (Hóa học sinh học của các nguyên tố: Hóa học vô cơ của sự sống.) (Nhà xuất bản Clarendon, 1991), ISBN 0-19-855598-9
- Nicholls, D. G. và Ferguson, S. J ., Bioenergetics. (Năng lượng sinh học.) (Academic Press Inc., 2002), ISBN 0-12-518121-3
Liên kết ngoài[sửa|sửa mã nguồn]
Thông tin chung
Trao đổi chất ở người
- Titles in NetBiochem Chủ đề trong hóa sinh ở người – Chỉ dẫn trong trao đổi chất ở người. Trình độ học sinh
- The Medical Biochemistry Page (Trang hóa sinh ở người) Nguồn tài nguyên toàn diện để hiểu quá trình chuyển hóa ở người.
Kho dữ liệu
Con đường chuyển hóa
Xem thêm: Điện Lạnh Duy Tân
Source: https://thevesta.vn
Category: Dịch Vụ